
- 果胶多糖结构与降血脂研究进展
- 发布时间: 2019-02-20 点击次数:5261
-
果胶多糖结构与降血脂研究进展
蔡为荣1 ,孙元琳2 ,汤 坚3
(1.安徽工程科技学院生化系,安徽 芜湖 241000;2.山西运城学院生命科学系,山西 运城 044000;
3. 江南大学 食品科学与技术国家重点实验室,江苏 无锡 214122)
专题论述 食品科学 2010, Vol. 31, No. 05
摘 要:果胶是一种广泛存在于植物细胞壁中的多糖类大分子物质,来源丰富,无毒,是重要的食品添加剂之一。随着高血脂人群的增多,果胶多糖调节血脂的功能结构日益受到关注。本文综述果胶多糖的结构、胆固醇吸收、 胆汁酸的束缚与排出、脂蛋白代谢变化及结肠酵解的研究进展,为果胶多糖降血脂生物活性及构效关系的进一步研究提供参考。
关键词:果胶多糖;结构;降血脂;研究进展
果胶是以原果胶、果胶、果胶酸的形态广泛分布于植物的果实、根、茎、叶中的多糖类高分子化合物, 与纤维素、半纤维素相伴,构成植物细胞胞间层(middlelamella)和初生壁(primary wall)。随着人们生活水平的日益提高,肥胖、高血压以及高脂血症等一些所谓“富贵病”的发病率呈逐年上升之势,及随功能性多糖的开发研究,果胶多糖及其降血脂功能的研究越来越受到研究和加工行业的重视。果胶多糖结构复杂,其生理活性与其结构紧密相关,各国学者进行了大量的研究工作,本文就果胶多糖结构与调节血脂功能作一综述。
1 结构和化学组成
就化学组成和生物合成而论,果胶可能是最复杂的一类多糖。果胶类多糖代表高等植物初级细胞壁和相邻
细胞间紧密联合的一组多糖,也代表从植物材料制备的一类复杂胶状多聚体,习惯称为果胶糖物质。其化学结构和分子量与其植物来源、组织部位、成熟度及提取方法等因素有关。早在 1825 年,Raconnot 首次从胡萝卜中提取出一种水溶性物质,可形成凝胶,并将该物质命名为“Pectin”(pectin 源于希腊词pektos,有凝固、凝结之意) [1]。1944 年 4 月美国化学会正式公布“果胶类物质的修订名称”,对当时常用的果胶酸( p e c t i n
acid)、果胶(pectin)和原果胶(protopectin)名词分别规定了聚半乳酸醛酸、含有中性糖侧链且部分被甲酯化的果胶酸、与纤维素和半纤维素结合在一起的甲酯化聚半乳糖醛酸苷的含义。随着科学技术的发展,众多学者在果胶多糖分离纯化、甲基化分析碳链结构和酸或酶水解等方面做了众多研究。如,1955 年Bishop[ 2 ] 报道:采用草酸铵缓冲液提取向日葵盘多糖,酸化乙醇与稀碱醇沉分别制得其果胶与多糖。纸层析实验显示,该果胶基本结构为α(1 → 4)键合的D- 半乳糖醛酸,多糖由半乳糖、葡萄糖和少量阿拉伯糖构成。1965 年,Zitko 等[ 3 ] 从向日葵、甜菜、苹果和柑橘分离出果胶酸,电泳实验显示这些果胶酸均由游离聚半乳糖醛酸糖和支链键合聚半乳糖醛酸糖两部分构成。发现在苹果[ 4 ] 、苜蓿[ 5 ] 、豆荚[6]中由半乳糖醛酸(1→2)鼠李糖构成的聚鼠李半乳糖醛酸含量较高。1973 年,Talmadge 等[ 7] 从无花果皮提取纯化聚鼠李半乳糖醛酸,为避免酸对糖苷键有不同程度的水解、过高的碱浓度会由于发生脱酯和β - 消去反应的影响,采用内切聚半乳糖醛酸酶水解,释放出各种多糖片段,甲基化后单糖醛醇乙酸酯质谱分析表明: 鼠李半乳糖醛酸聚糖主要呈鼠李糖-(1 → 4)- 半乳糖醛酸(1 → 2)鼠李糖碳链结构。1978 年,Darvill 等[8]用高纯的内切聚半乳糖醛酸酶水解 7 种悬铃木细胞,凝胶色柱纯化得到的鼠李半乳糖醛酸聚糖 - Ⅱ - ( R h a m n o - galacturonans II,RG- Ⅱ)含量相近,RG- Ⅱ由半乳糖醛酸(GalA)、鼠李糖(Rha)、阿拉伯糖(Ara)、半乳糖(Gal) 等 10 种糖残基组成;鼠李糖残基以 2 ,4 糖苷键连接在D- 半乳糖醛酸残基形成主链;侧链上主要含 L- 阿拉伯糖和 D- 半乳糖残基。1997 年,Renard 等[9 ]进一步应用HPLC 色谱和1 H NMR 分析了各种多糖碎片,证实了以上推测。一个半世纪以来,众多科学家在果胶多糖领域进行着细致而深入的研究,使果胶多糖的结构、化学组成越来越趋于系统和完善。
果胶多糖的分子量介于 10000~400000D 之间,其分子结构分为光滑区与毛发区:光滑区由α -D- 半乳糖醛酸残基通过 1,4 糖苷键线形连接;毛发区由高度分支的α - L - 鼠李半乳糖醛酸组成。因此,按照结构分类, 果 胶类多糖包括 3 大类 : 半乳糖醛酸聚糖 (homogalacturonans,HGA)、鼠李半乳糖醛酸聚糖- Ⅰ(rhamnogalacturonans I,RG- Ⅰ)和鼠李半乳糖醛酸聚糖- Ⅱ。
1.1 半乳糖醛酸聚糖(HGA)
HGA是由α-D- 半乳糖醛酸残基通过1,4 糖苷键连接而成的线性糖链,由 100~500 个GalA 残基组成。其中半乳糖醛酸的羧基有不同程度的甲酯化,因此天然果胶分为高甲氧基果胶(HM)和低甲氧基果胶(LM)。未甲酯化的羧基则以游离酸形式或以K、Na、Ca 盐等形式存在。HM 果胶在有足够的糖浓度和酸条件下才能形成凝胶;LM 果胶在Ca2+ 存在时通过半乳糖醛酸链与Ca2+ 交联发生分子间缔合而形成凝胶(图 1)[10]。有的GalA 的C2 或C 3 位羟基还发生乙酰化。
1.2 鼠李半乳糖醛酸聚糖- Ⅰ(RG- Ⅰ)
果胶多糖通常为杂多糖,常在α -(1 → 4)连接的多聚半乳糖醛酸主链中间插入吡喃型鼠李糖(Rhap),形成半乳糖醛酸和鼠李糖交替组成的重复单位:4 )- α - D - GalpA-(1,2)- α -L-Rhap-(1,即为RG- Ⅰ(图 2)[11]。在鼠李糖残基的C4 位结合有不同种类的中性或酸性寡聚糖:
1)阿拉伯半乳聚糖(arabinogalactans)以β -(1 → 4)连接的多聚半乳糖为主链并带有α -(1 → 5)L- 阿拉伯呋喃糖残基(Araf)侧链;2)半乳聚糖(galactan)是高度分支的多糖,
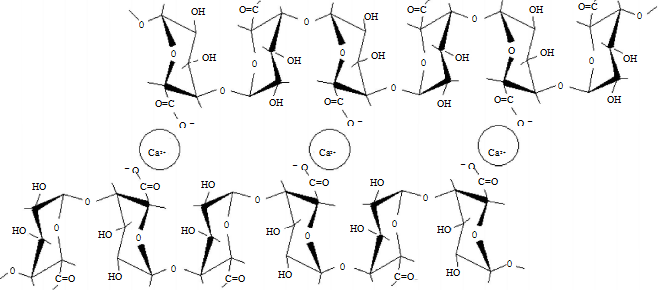
图 1 LM“蛋盒”胶凝模型
Fig.1 Schematic egg box model of calcium binding to low-methoxyl pectin
由β -D- 吡喃半乳糖残基以 1,3 和 1,6 键相互连接而成;
3)阿拉伯聚糖(arabinan)由 L- 阿拉伯糖残基以 1,5 键连接, 并接有 1 , 3 分支。此外,毛发区的侧链末端还发现有α -L- 果糖残基(Fucp),β -D- 葡萄糖醛酸残基(GlcpA), 以及4-O- 甲基- β-D- 葡萄糖醛酸残基(4-O-Me GlcpA)等。
1.3 鼠李半乳糖醛酸聚糖-II(RG-II)
RG-II是所有高等植物的细胞壁中均含有的一种酸性果胶,其结构特征是主链为均聚半乳糖醛酸,侧链含有 4 种结构复杂的寡糖(图 3)。与细胞壁中的硼形成复合物,参与硼的运输[12]。RG-II 多糖能与铅离子鳌合,具有解毒与免疫调节活性[13]。Nergard 等[14] 研究表明,从斑鸠菊的根中提取的含有RG- II 结构的酸性果胶物质, 具有补体修复以及促进B 与T 细胞生长活性。
2 果胶多糖调节血脂机理
早在 20 世纪 60 年代,在人和大鼠的研究中就已发现果胶多糖能降低血清胆固醇水平[16- 17 ],从此,在胆固醇的消化、吸收、转运等方面开展了有关果胶多糖降低血清胆固醇的研究。
2.1 果胶多糖对胆固醇吸收的影响
机体内的胆固醇主要有两种来源,一种是肠道内吸收的膳食胆固醇、胆固醇酯和胆汁中的胆固醇,另一种是肝脏和外周组织中产生的胆固醇。饮食中胆固醇酯在胰腺分泌的胆固醇酯酶水解下生成胆固醇,与胆汁酸、磷脂、甘油脂一起组成混合微团,吸入黏膜细胞, 在内质网内形成乳糜微粒,经淋巴系统进入血液完成消化吸收。果胶吸水性强且难消化,从而可充盈肠胃。但 对胰腺分泌仅有很小影响[18 ]。Jacobs[19 ]认为果胶会增加绒毛细胞的脱落。Kelley 等[20]采用14C 标记胆固醇,考察结果显示,摄入果胶、瓜尔胶、车前子胶均可使肝中胆固醇吸收下降,仅果胶可使血浆胆固醇吸收下降。多数学者认为,果胶具有凝胶特性,影响胆固醇与消化酶、胆汁酸微团及肠黏膜的接触,同时束缚胆酸,造成脂质乳化障碍[ 2 1 ] ,减慢肠系膜的淋巴循环及胆汁再循环。
2.2 果胶多糖对胆汁酸的束缚与排出的影响
胆汁酸在肝脏中由胆固醇合成后经肠肝循环排入十二指肠,维持人体对脂质物质的消化吸收。正常情况下,大部分胆汁酸在小肠中经肝肠循环重吸收进入肝脏,从而使胆汁酸保持稳定。果胶束缚胆汁酸、促进其排出,通过肝内胆固醇池平衡,调用血浆胆固醇, 从而起到降胆固醇的作用。
1990 年,美国Fernandez 等[22]报道,胆固醇饲料加1% 仙人掌果胶,喂养几内亚猪 25d,血浆低密度脂蛋白胆固醇(LDL-C)下降 33%,肝细胞微粒体的 3- 羟基-3- 甲基戊二酰辅酶A 还原酶(胆固醇合成限速酶)浓度没有变化,游离胆固醇与胆固醇酯分别下降 40 % 与 8 5 %。添加果胶( 1 2 . 5 %) 到低、高胆固醇饲料中,可使肝内7α - 羟化酶(胆汁酸合成限速酶)活性分别上调3倍和5倍[23]。结果显示果胶下调血浆胆固醇的途径,类似于胆汁酸结合树脂。
果胶对脂质代谢的作用,与其结构、黏度、阳离子交换特性及消化道内的分泌物变化有关。Terpstra 等报道:添加高黏度钙敏感的果胶( DE,69 %)、低黏度钙不敏感的果胶( DE,34 %)、高黏度瓜尔豆胶、低黏度瓜尔豆胶和车前子胶于饲料中,大鼠 8 周喂养后结果显示: 高黏度果胶实验鼠中血浆胆固醇酯转运蛋白(CETP)活性比低黏度果胶显著下降;粪便中胆汁酸和中性甾醇增多[ 24 ];而添加 8% 车前子胶,肝中总胆固醇和胆固醇酯比果胶组有显著下降,粪便量增加 9 0 %,胆(酸) 盐、鹅脱氧胆( 酸) 盐、甘氨酸和牛磺酸含量显著提高[ 25 ] 。提示果胶与车前子胶降血脂机制有不同之处。
2.3 果胶多糖对脂蛋白代谢调控的影响
ABCA1 和ATP 为结合转运蛋白;ACAT 为脂肪酰- 胆固醇酰基转移酶;CETP 为胆固醇酯转运蛋白; CE 为胆固醇酯; CM 为糜乳微粒;CR 为残余颗粒;HL 为肝脂肪酶;LDL-R 为LDL 受体;Ch 为胆固醇; SR-B1 为清道夫受体-B1。
图 4 胆固醇及载脂蛋白转运示意图
Fig.4 Schematic diagram of cholesterol and lipoprotein metabo- lism
20 世纪初大量的流行病学研究发现血浆胆固醇(TC) 水平、甘油三酯(TG) 与冠心病的病死率密切相关。90 年代通过对脂质,尤其是脂蛋白的研究显示, LDL-C 增高促进动脉粥样硬化,高密度脂蛋白- 胆固醇(HDL-C) 增高则有防止动脉粥样硬化作用。HDL- C 视为“好的胆固醇 ”。
Fernandez 等[22 ]研究报道,摄入果胶、瓜尔胶、车前子胶后,血浆胆固醇显著下降;极低脂蛋白(VLDL) 中TG 和磷脂含量增加,暗示有新生的VLDL 生成;卵磷脂胆固醇脂酰转移酶(LCAT)和CETP 活性下降;放射性标记VLDL测得VLDL转运更快、LDL-apoB 外流提高, 表明果胶等物质的摄入可上调LDL 受体。Brown 等[26]进一步研究指出,摄入每克果胶可分别下调总胆固醇和低密度脂蛋白胆固醇约 0.070mmol/L 和 0.055mmol/L。
近代医学研究显示,清道夫受体-B1(SR-B1)和腺三磷结合盒转运体A1(ABCA1)在介导细胞内胆固醇流出中发挥重要作用[27](图4)。SR-B1 介导的肝脏可从HDL 选择性摄取胆固醇。其中apo A Ⅰ是 HDL 最主要的蛋白成分,与高密度脂蛋白颗粒的形成有关;是卵磷脂胆固醇酰基转移酶(LCAT)的主要激活剂。HDL 可将蓄积于末梢组织的游离胆固醇与血循环中的脂蛋白或某些大分子结合,运输到肝脏,进行胆固醇的逆转运。A B C A 1 是载脂蛋白介导的脂质逆转运通路中的限速蛋白质,转录后的调节可通过与apoA Ⅰ的结合来实现。现已知胆固醇、acLDL、oxLDL 等多个因素可以调节 ABCA1 的活性[ 2 8 ] 。
李茂言等[29]研究了荞秸果胶对高脂血症大鼠调脂作用,结果显示荞秸果胶呈剂量依赖性地明显降低肝胆固醇、TC、LDL-C,高密度脂蛋白亚组分3 胆固醇(HDL3-C) 及血清TG 的含量。可明显提高高密度脂蛋白亚组分2 胆固醇(HDL2-C) 的含量。
此外,瓜尔豆胶、甜菜多糖可上调LDL 受体和下调SR-B1 活性[30-31]。
2.4 果胶多糖对结肠发酵作用的影响
机体本身不分泌降解果胶的酶,果胶可被大肠内的 微生物发酵降解,产生短链脂肪酸( SCFA),后经门脉系统进入肝脏代谢。Gulfi 等报道果胶的分子量和黏度并不影响其酵解性,高甲氧基果胶产生的SCFA 量稍多, 高甲氧基果胶完全酵解时间短,低甲氧基果胶完全酵解时间长[32 ],肠菌对果胶代谢的主要终末产物是乙酸、丙酸、丁酸、二氧化碳、甲烷、氢气和水。可溶性纤维果胶生成乙酸、丙酸、丁酸的比例约为 80 : 12 : 8 [ 3 3 ] 。乙酸、丙酸、丁酸被结肠黏膜上皮细胞吸收。果胶毛发区生成丙酸比例比阿拉伯胶、低甲氧基苹果果胶、高甲氧基苹果果胶高[34 ]。动物实验表明,丙酸盐可使血浆
胆固醇水平下降[35];乙酸盐抑制肝内胆固醇的合成,并
使血浆胆固醇向肝内转移[36]。也有人认为酵解增加了大肠内细菌的活性,使胆汁酸离散,不易被结肠黏膜吸收而被排出,导致从头合成胆固醇来补充胆汁酸[ 3 7 ] 。
综上所述,果胶多糖是通过降低胆固醇吸收、促进胆汁酸的束缚与排出、上调LD L 受体、加速载脂蛋白转运和结肠发酵作用,多途径调控机体内胆固醇及甘油三酯含量。由于果胶多糖结构复杂性,胶凝性、结肠发酵程度均不同。其产物对胆固醇代谢、残渣对胆汁酸束缚、以及机体肠道菌、益生菌与肠道蠕动等诸多因素相互影响,果胶多糖降血脂的量效关系尚需进一步研究。
3 结 语
近 20 年来,伴随着生活节奏的加快、饮食结构的改变,人们体力活动的减少,高血脂症成为威胁人类生命健康的常见病、多发病。我国调节血脂保健食品市场曾一度混乱,良莠不齐。确定功能因子和构效、量效关系的第二产品较少。果胶多糖资源丰富,对机体无副作用,在食品及医药工业应用广泛。果胶多糖降血脂机理侧重于胆汁酸的束缚与排出,进一步开展果胶多糖结构与作用机理的研究,可为降血脂多糖的开发提供参考。
Research Progress in Structure and Hypolipidmic Mechanisms of Pectic Polysaccharides
CAI Wei-rong1,SUN Yuan-lin2,TANG Jian3
(1. Department of Biochemistry, Anhui College of Technology and Science, Wuhu 241000, China;
2. Department of Life Sciences, Yuncheng University, Yuncheng 044000, China ;
3. State Key Laboratory of Food Science and Technology, Jiangnan University, Wuxi 214122, China)
Abstract :Pectin is complex polysaccharides in cell walls of all plants. It is one of the most important food additives due to its abundant source and safety. Increasing attention has been paid to using pectic polysaccharides for the hypolipidemic function because of increasing number of hyperlipidemic patients. In this paper, the structure of pectic polysaccharides, and the effects of pectic polysaccharides on cholesterol absorption, bile acid chelation and secretion, plasma lipoprotein cholesterol metabolism and colon glycolysis are reviewed. All of these investigations will provide a reference to further explore structure-activity relationship of pectic polysaccharides on hypolipidemic function.
Key words:pectic polysaccharide;structure;antihyperlipidemia;research progress
中图分类号: O636.1 文献标识码:A 文章编号:1002-6630(2010)05-0307-05
- 上一篇:低分子柑桔果胶粉对糖尿病小鼠血糖、血脂的调节作用
下一篇: - 返回新闻列表
